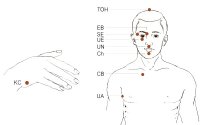
Biološka složenost ljudi
Velika složenost organa ljudskog tijela prkosi evolucijskom objašnjenju.
Darvinisti nisu detaljno objasnili kako su se ti organi razvili nasumičnim genskim varijacijama i prirodnom selekcijom.
Oko
Ljudsko oko jedan je od takvih naizgled neizvodivo složenih organa. Svjetlo prolazi kroz zjenicu, a leća fokusira svjetlo na mrežnicu. Oko posjeduje i svojstva koja ispravljaju smetnje između svjetlosnih valova različitih frekvencija. Oko ne može funkcionirati bez svih svojih sastavnih dijelova.
Čak je i Darwin uvidio da oko, kao i druge složene strukture, predstavljaju problem njegovoj teoriji evolucije, koja pretpostavlja da se takve strukture postupno razvijaju tijekom više naraštaja. Darwin nije podrobno objasnio
kako se to događa, no ukazao je na različita živa bića s različitim vrstama očiju - od kojih su neke samo fotosenzibilne točke, neke, pak, jednostavne udubine s jednostavnim lećama, a druge mnogo složenije. Pretpostavio je i da se ljudsko oko možda razvilo slijedom takvih etapa. Pritom je zanemario pitanje kako je nastala prva fotosenzibilna točka. "Pitanje kako živac postaje fotosenzibilan za nas nije toliko važno koliko pitanje postanka samoga života." (Darwin, 1872., str. 151; Behe, 1996., str. 16-18).
Darwinovo neodređeno objašnjenje načina na koji se fotosenzibilna točka postupno razvila u složeno ljudsko oko nalik kameri, možda je samo površno uvjerljivo, no to nije i znanstveno objašnjenje podrijetla oka. Ono je jednostavno poziv da zamislimo da se evolucija uistinu dogodila. Želimo li pretvoriti maštu u znanost, valja nam razmotriti građu oka na biomolekularnoj razini.
Devlin (1992., str. 938-954) prilično opsežno opisuje biokemijski proces razvoja ljudskog vida. Biokemičar Michael Behe (1996., str. 18-21) na sljedeći način sažima Devlinovo objašnjenje: "Kada svjetlo prvi put udari o mrežnicu, dolazi do reakcije fotona i molekule 11-
cis-retinal, koja se u pikosekundama preraspoređuje u trans-retinal... Promjena oblika retinal - ne molekule izaziva promjenu oblika bjelančevine rodopsin, uz koju je retinal usko povezan ... Bjelančevina, koja se danas naziva metarodopsin II, spojena je uz drugu bjelančevinu zvanu transducin. Prije sudara s metardopsinom II, transducin je čvrsto povezao malu molekulu GDP. No, kada transducin stupi u reakciju s metarodopsinom II, GDP ispada i molekula GTP se veže uz transducin ... GTP-transducin-metarodopsin II tada se veže uz bjelančevinu fosfordiesteraza, bjelančevinu smještenu u unutrašnjoj staničnoj opni. Vezujući se uz metarodopsin II i njezinu pratnju, fosfordiester stječe kemijsku sposobnost da izreže molekulu cGMP ...
Stanica izvorno sadrži velik broj molekula cGMP, no fosfordiester smanjuje tu koncentraciju, kao što se uslijed izvlačenja čepa snižava razina vode u kadi. Druga opnasta bjelančevina koja veže cGMP zove se ionski kanal.
On djeluje kao prolaz koji regulira količinu natrijevih iona u stanici, dok ih zasebna bjelančevina iznova izbacuje. Istodobno djelovanje ionskog kanala i pumpe održava razinu natrijevih iona u stanici u ograničenom omjeru.
Kada se količina cGMP smanji zbog cijepanja djelovanjem fosfordiestera, ionski kanal se zatvara, ujedno smanjujući koncentraciju pozitivno nabijenih natrijevih iona u stanici. To uzrokuje neuravnoteženost naboja preko stanične opne, što pak, naposljetku uzrokuje prijenos struje niz optički živac do mozga. Mozak tu posljedicu tumači kao vid."
Drugi jednako složen niz reakcija obnavlja izvorne kemijske elemente koji su započeli taj proces, kao što su 11-
cis-retinal, cGMP i natrijevi ioni (Behe, 1996., str. 21). A to je samo dio biokemije koja se odvija u vizualnom procesu. Behe je izjavio (1996., str. 22): "U konačnici... to je razina objašnjenja kojoj biološka znanost mora stremiti. Da bismo mogli shvatiti funkciju, valja nam razumjeti svaku značajnu etapu tog procesa. Značajne se etape bioloških procesa u konačnici događaju na molekularnoj razini, pa zadovoljavajuće objašnjenje biološkog fenomena - kao što su vid, probava ili imunitet - mora uključivati njegovo molekularno objašnjenje."
Evolucionisti nisu iznijeli takvo objašnjenje.
Mjehuričasti prijenosni sustav
Lizosom je odjeljak (stanična organela) u citoplazmi stanice koji se rješava oštećenih bjelančevina. U lizosomu postoje enzimi koji rastavljaju bjelančevine.
Ti se enzimi proizvode u ribosomima, odjeljcima u sklopu drugog staničnog odjeljka (organele) zvanog endoplazmatska mrežica. Budući da enzimi nastaju u ribosomima, obilježeni su posebnim redoslijedom aminokiselina, koji im dopušta prolaz kroz zidove ribosoma u endoplazmatsku mrežicu. Ondje bivaju obilježeni drugim slijedom aminokiselina, koji im dopušta da izađu iz endoplazmatske mrežice. Enzimi putuju do lizosoma, gdje se vežu za njegovu površinu. Potom im drugi niz signalnih obilježja dopušta da uđu u lizosom, gdje mogu izvršiti svoju funkciju (Behe, 1998., str. 181-182; Alberts i dr., 1994., str. 551-650). Ta prijenosna mreža naziva se mjehuričastim prijenosnim sustavom.
U slučaju bolesti stanice, pogreška u signalnom označavanju ometa mjehuričasti prijenosni sustav. Umjesto da prenosi enzime koji sadrže raspadajuće bjelančevine iz ribosoma u lizosome, taj ih sustav prenosi do staničnog zida, odakle se izbacuju iz stanice. U međuvremenu, oštećene bjelančevine teku u lizosome, gdje se ne raspadaju. Lizosomi se bez enzima koji sadrže raspadajuće bjelančevine, pune poput prenatrpanih kanti smeća. Da bi to spriječila, stanica proizvodi nove lizosome, koji se također ispunjavaju otpadnim bjelančevinama. Naposljetku, kada je velik broj lizosoma ispunjen otpadnim bjelančevinama, čitava se stanica raspada i pacijent koji boluje od spomenute bolesti umire. To pokazuje što se događa kada nedostaje jedan dio složenog sustava - raspada se čitav sustav. Da bi mjehuričasti prijenosni sustav mogao učinkovito djelovati, svi njegovi dijelovi moraju se nalaziti na svojemu mjestu.
Behe kaže (1996., str. 115-116): "Mjehuričasti prijenos uistinu je zbunjujući proces, koji se po složenosti može usporediti s automatskim dostavljanjem cjepiva iz spremišta u bolnicu udaljenu više tisuća kilometara.
Poremećaji u mjehuričastom prijenosu mogu imati iste kobne posljedice kao i pogreške u dostavljanju neophodnog cjepiva gradu pogođenom pandemijom. Analiza pokazuje da je mjehuričasti prijenos veoma složen, radi čega njegov razvoj prkosi objašnjenjima o postupnosti na kojima ustrajava darvinistička evolucija. Pregledom stručne biokemijske literature, uviđa se da nitko nikada nije pokušao detaljno opisati put nastanka takvog sustava. S obzirom na izvanrednu složenost mjehuričastog prijenosa, darvinistička teorija ostaje bez objašnjenja."
Mehanizam zgrušavanja krvi
Ljudski mehanizam zgrušavanja krvi za evolucioniste predstavlja drugu zagonetku. Behe kaže (1996., str. 78): "Zgrušavanje krvi izrazito je složen, zamršeno isprepleten sustav sastavljen od mnoštva međusobno zavisnih dijelova bjelančevina. Nedostatak bilo koje od brojnih sastavnica, ili bitni nedostaci u njima, uzrokuju raspad sustava: krv se ne zgrušava u prikladno vrijeme ili na prikladnome mjestu." Stoga je taj sustav nepromjenjivo složen i ne može se jednostavno objasniti stupnjevima darvinističke evolucije.
Mehanizam zgrušavanja krvi osniva se na fibrinogenu, krvnoj bjelančevini koja oblikuje tkiva koja tvore ugruške. Fibrinogen se obično rastvara u krvnoj plazmi. Kada počne krvarenje, bjelančevina trombin siječe fibrinogen i tako stvara nizove bjelančevine fibrin. Vlakna fibrina su međusobno spojena i tvore mrežu koja hvata krvne stanice, zaustavljajući, tako, istjecanje krvi iz rane (Behe, 1996., str. 80). Mreža prvotno nije veoma čvrsta.
Katkad se slama, uslijed čega krv iznova istječe iz rane. Da bi se to spriječilo, bjelančevina koja se zove faktor stabilizacije fibrina (FSF) isprepliće vlakna fibrina, čime osnažuje mrežu (Behe, 1996., str. 88).
U međuvremenu, trombin nastavlja sjeći fibrinogen u fibrine, koji tvore dodatne ugruške. U određenom trenutku, trombin mora prestati rezati fibrinogen, jer će u suprotnom biti proizvedena prevelika količina fibrina koji će zgrušati čitav krvni sustav, što dovodi do smrti čovjeka (Behe, 1996., str. 81).
Postoji čitavo mnoštvo bjelančevina i enzima koji sudjeluju u uključivanju i isključivanju sustava zgrušavanja krvi u odgovarajuće vrijeme. Trombin prvotno postoji u nedjelatnom obliku protrombina. U tom obliku on ne siječe fibrinogen u vlakna fibrina koja tvore ugruške. Prema tome, da bi proces zgrušavanja mogao početi, protrombin mora biti pretvoren u trombin. U suprotnom će čovjek iskrvariti do smrti. A kada se oblikuje odgovarajući sustav zgrušavanja, trombin se mora iznova pretvoriti u protrombin.
U suprotnom će se zgrušavanje nastaviti sve dok krv ne prestane istjecati (Behe, 1996., str. 82).
U aktivaciji protrombina sudjeluje bjelančevina koja se naziva Stuartov faktor, a koja ga pretvara u trombin, čime omogućuje početak procesa zgrušavanja. Što, dakle, potiče taj pasivan Stuartov faktor? Postoje dva niza interakcija, koje počinju preobrazbama na području rane. Razmotrimo samo jednu od njih. Behe kaže (1996., str. 84): "Kod životinje prerezanog tijela, bjelančevina HMK cijepa bjelančevinu koja se zove Hagemanov faktor, uslijed čega nastaje aktivan Hagemanov faktor. Aktivirani Hagemaov faktor odmah pretvara drugu bjelančevinu, prekalikrein u njegov aktivan oblik, kalikrein. Kalikrein pomaže HMK u ubrzavanju pretvaranja Hagemanova faktora u njegov aktivan oblik. Aktivirani Hagemanov faktor i HMK potom zajedno pretvaraju drugu bjelančevinu, PTA, u njezin aktivan oblik. Aktivirana PTA, pak, zajedno s aktiviranim oblikom druge bjelančevine zvane konvertin, pretvara bjelančevinu zvanu Christmas faktor u njezin aktivan oblik. Konačno, aktivirani Christmas faktor, zajedno s antihemofilijskim faktorom ... pretvara Stuartov faktor u njegov aktivan oblik."
Drugi je niz jednako složen i na nekim se mjestima pretapa s prvim.
Dakle, sada imamo aktiviran Stuartov faktor. Ali čak ni to nije dovoljno za početak procesa zgrušavanja. Prije nego li Stuartov faktor može početi djelovati na protrombin, potonji se mora transformirati promjenom svojih deset podjedinica aminokiselina. Nakon tih promjena, protrombin se može prilijepiti uz rub stanice. Protrombin se može promijeniti (djelovanjem Stuartova faktora) u trombin koji potiče zgrušavanje, tek nakon što je spojen uz rub stanice. Lijepljenje protrombina uz rub stanice u blizini rane, pomaže usmjeravanju procesa zgrušavanja točno na područje rane. No, aktiviran Stuartov faktor veoma sporo pretvara protrombin u trombin. Organizam umire prije stvaranja dovoljne količine trombina, koja će omogućiti početak učinkovitog procesa zgrušavanja. Stoga je neophodna prisutnost druge bjelančevine, akcelerina, koji ubrzava djelovanje Stuartova faktora na protrombin (Behe, 1996., str. 81-83).
Protrombin je, dakle, sada pretvoren u trombin. Trombin cijepa fibrinogen, stvarajući fibrin koji oblikuje ugruške. Sada se možemo osvrnuti na pitanje kako se taj proces zgrušavanja može zaustaviti prije nego li započne.
Neusmjereno zgrušavanje može začepiti krvne žile u organizmu, što može imati kobne posljedice. Nakon oblikovanja molekula trombina, uz njih se veže bjelančevina antitrombin, koja ih, tako, deaktivira. Ali antitrombin se veže samo u doticaju s drugom bjelančevinom, heparinom, koja se nalazi u neoštećenim krvnim žilama. To, dakle, znači da se antitrombin veže uz aktivirane molekule trombina tek nakon njihova ulaska u neoštećene krvne žile, čime ih deaktivira i zaustavlja zgrušavanje. Zgrušavanje se može nastaviti u oštećenoj krvnoj žili. Na taj se način zgrušavanje nastavlja samo u području rane, a ne u drugim neoštećenim krvnim žilama.
Nakon što se oštećena žila oporavi, na tom će mjestu prestati i zgrušavanje.
Do toga dolazi uslijed složenog procesa, kao što je proces zaustavljanja zgrušavanja krvi u neoštećenim krvnim žilama (Behe, 1996., str. 87-88).
Nakon proteka određenog razdoblja, kada rana zacijeli, valja ukloniti sam ugrušak. Bjelančevina plazmin reže mrežu fibrina koja tvori ugrušak.
Kako se dade zaključiti, plazmin izvorno postoji u pasivnom obliku plazminogena, i mora se aktivirati u odgovarajućem trenutku kako bi mogao ukloniti ugrušak. Naravno, njegova aktivacija uključuje složene interakcije s drugim bjelančevinama (Behe, 1996., str. 88).
Behe kaže (1996., str. 86): "Sustav zgrušavanja krvi s pravom se može opisati kao nepromjenjivo složen sustav. Drugim riječima, to je sustav sastavljen od nekoliko interaktivnih dijelova koji omogućavaju osnovnu funkciju, dok uklanjanje samo jednog od tih dijelova uzrokuje potpun prestanak funkcioniranja sustava ... U nedostatku samo jedne od tih sastavnica, krv se ne zgrušava i sustav propada. Evolucionisti nisu pružili zadovoljavajuće objašnjenje na koji je način nastao taj složen kemijski sustav održavanja, koji sadrži brojne jedinstvene bjelančevine s veoma određenim funkcijama.
Stručnjak za zgrušavanje krvi, Russell Doolittle, jednostavno tvrdi da su neophodne bjelančevine u tom sustavu rezultat duplikacije i miješanja gena. No, procesom duplikacije gena proizvodi se samo duplikat već postojećeg gena. Doolittle ne specificira koje su mutacije tog dupliciranog gena neophodne da bi bjelančevini koju proizvodi, dao novu funkciju korisnu u nekom razvojnom sustavu zgrušavanja. Miješanje gena osniva se na ideji da je svaki gen sastavljen od nekoliko podiedinica. Katkad se u procesu reprodukcije dijelovi gena odvajaju i međusobno spajaju u novom rasporedu.
Iznova izmiješan gen proizvodi drukčiju bjelančevinu. Ali, vjerojatnost da se odgovarajuća količina podjedinica gena spoji i oblikuje nov gen, koji bi proizveo bjelančevinu korisnu za proces zgrušavanja krvi, gotovo je nemoguća. Jedna bjelančevina u sustavu, TPA, sastoji se od četiri dijela. Pretpostavimo da je u vrijeme kada se tek počeo oblikovati sustav zgrušavanja krvi i kada nije bilo TPA, postojala životinja. Nadalje, pretpostavimo da je ta životinja imala 10.000 gena. Svaki je gen u prosjeku podijeljen na tri podjedinice. To čini dostupnim 30.000 gena za miješanje gena.
Vjerojatnost da će se četiri dijela koja tvore TPA, nasumce spojiti je 30.000
4 - dakle, veoma je mala. No, glavni problem predstavlja međusobno spajanje svih dijelova u djelatni sustav. Samo na takav sustav, koji pridonosi zdravlju organizma može djelovati prirodna selekcija. Izolirani dijelovi sustava ustvari ne pridonose zdravlju, i stoga nije moguća prirodna selekcija.
Dakle, u nastojanju da objasne postojanje sustava zgrušavanja u ljudskom tijelu, evolucionisti su najprije morali dokazati postojanje jednostavnog sustava zgrušavanja krvi i pokazati, detaljno, kako promjene u genima mogu proizvesti sve veći broj učinkovitih sustava koji pridonose zdravlju organizma. No, to nisu učinili (Behe, 1996., str. 90-97). Neki su znanstvenici, nastojeći izbjeći te kritike, pretpostavili da su dijelovi takvog složenog sustava možda imali druge funkcije u drugim sustavima prije nego li su se spojili u spomenutom sustavu. No, to dodatno otežava već zamršeno pitanje. U spomenutom slučaju, znanstvenici bi trebali pokazati kako su ti drugi sustavi s različitim funkcijama nastali postupno, i kako su dijelovi njihovih sustava kooptirani s drugom svrhom, bez oštećivanja tih sustava.
















































 bglavac
bglavac vanessa
vanessa irida
irida